GtoPdb is requesting financial support from commercial users. Please see our sustainability page for more information.

 Target has curated data in GtoImmuPdb
Target has curated data in GtoImmuPdb
Contents:
Gene and Protein Information  |
||||||
| Species | TM | AA | Chromosomal Location | Gene Symbol | Gene Name | Reference |
| Human | 1 | 824 | 2p25.1 | ADAM17 | ADAM metallopeptidase domain 17 | |
| Mouse | 1 | 827 | 12 8.3 cM | Adam17 | a disintegrin and metallopeptidase domain 17 | |
| Rat | 1 | 827 | 6q16 | Adam17 | ADAM metallopeptidase domain 17 | |
| Previous and Unofficial Names |
| CD156B | cSVP | snake venom-like protease | TACE | TNF α convertase | TNF α-converting enzyme | tumor necrosis factor, alpha, converting enzyme |
| Database Links |
|
| Specialist databases | |
| MEROPS | M12.217 (Hs) |
| Other databases | |
| Alphafold | P78536 (Hs), Q9Z0F8 (Mm), Q9Z1K9 (Rn) |
| BRENDA | 3.4.24.86 |
| CATH/Gene3D | 4.10.70.10, 3.40.390.10 |
| ChEMBL Target | CHEMBL3706 (Hs), CHEMBL4379 (Mm), CHEMBL2523 (Rn) |
| Ensembl Gene | ENSG00000151694 (Hs), ENSMUSG00000052593 (Mm), ENSRNOG00000060694 (Rn) |
| Entrez Gene | 6868 (Hs), 11491 (Mm), 57027 (Rn) |
| Human Protein Atlas | ENSG00000151694 (Hs) |
| KEGG Enzyme | 3.4.24.86 |
| KEGG Gene | hsa:6868 (Hs), mmu:11491 (Mm), rno:57027 (Rn) |
| OMIM | 603639 (Hs) |
| Orphanet | ORPHA299456 (Hs) |
| Pharos | P78536 (Hs) |
| RefSeq Nucleotide | NM_003183 (Hs), NM_001277266 (Mm), NM_009615 (Mm), NM_020306 (Rn) |
| RefSeq Protein | NP_003174 (Hs), NP_033745 (Mm), NP_001264195 (Mm), NP_064702 (Rn) |
| SynPHARM | 82602 (in complex with IK-862) |
| UniProtKB | P78536 (Hs), Q9Z0F8 (Mm), Q9Z1K9 (Rn) |
| Wikipedia | ADAM17 (Hs) |
| Selected 3D Structures |
|||||||||||||
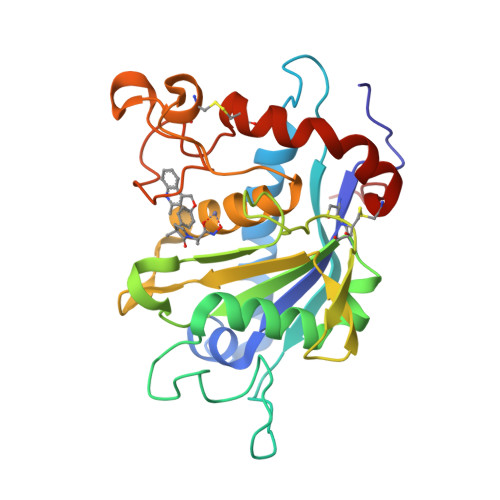
|
|
||||||||||||
| Enzyme Reaction |
||||
|
||||
Download all structure-activity data for this target as a CSV file 
| Inhibitors | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Key to terms and symbols | View all chemical structures | Click column headers to sort | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| View species-specific inhibitor tables | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||


| Antibody Comments | ||
| An anti-ADAM17 (TACE; TNFα converting enzyme) monoclonal antibody, MEDI3622 [11], has shown anti-proliferative activity in tumour models (IC50 ~26 nM vs. H292 cells) [12]. MEDI3622 binds ADAM17 with a Kd of 0.039 nM and inhibits its catalytic activity with a Ki of 3.1 nM [12]. |
| Immunopharmacology Comments |
| The proteolytic activity of ADAM17 (a type I transmembrane metalloproteinase; a.k.a.TNF-alpha converting enzyme or TACE) is involved in the shedding of the extracellular domains of several transmembrane proteins e.g. cytokines (TNFα), growth factors, receptors (IL-6R and TNF-R for example) and adhesion molecules. Cleavage of substrates, including TNFα, IL-6R and L-selectin, produce pro-inflammatory effects stimulating both innate and acquired immune responses. ADAM17 activity is crucial during development (ADAM17 knockout is embryonic lethal), and it has been shown that the soluble IL-6R/IL-6 complex generates agonist-like signals in a process termed IL-6 trans-signaling. The generation and maintenance of several inflammatory and autoimmune diseases is driven by IL-6 trans-signaling [1]. ADAM17 activity may also be involved in directing migration of activated lymphocytes and neutrophils in vivo, by catalysing the shedding of L-selectin from the surface of leukocytes. ADAM17 is being investigated as a novel therapeutic target in immune-related diseases such as rheumatoid arthritis (RA) [7], but also plays roles in tissue regeneration and cancer development. |
| Immuno Process Associations | ||||||||||||||||||
|
||||||||||||||||||
|
||||||||||||||||||
|
||||||||||||||||||
|
||||||||||||||||||
|
||||||||||||||||||
|
||||||||||||||||||
|
||||||||||||||||||
|
| Clinically-Relevant Mutations and Pathophysiology
|
||||||||||||
|
||||||||||||
| General Comments |
| There is evidence that ADAM17 cleaves angiotensin converting enzyme 2 (ACE2), leading to shedding of the ACE2 ectodmain as a soluble entity [4,9]. Given that ACE2 is the principal binding site for SARS-CoV-2 [13], it could be forseen that exploiting ADAM17's ACE2-sheddase activity (for example using a pharmacological activator, increasing its phosphorylation by upstream mediators, or otherwise upregulating its expression) could lead to release of ACE2/coronavirus complexes, as a mechanism to reduce the ability of the virus to infect lung epithelial cells. The obvious caveat to this approach would be to consider the effect that reducing membrane ACE2 could have on the renin-angiotensin system and other biological processes. |
References
1. Chalaris A, Garbers C, Rabe B, Rose-John S, Scheller J. (2011) The soluble Interleukin 6 receptor: generation and role in inflammation and cancer. Eur J Cell Biol, 90 (6-7): 484-94. [PMID:21145125]
2. DasGupta S, Murumkar PR, Giridhar R, Yadav MR. (2009) Current perspective of TACE inhibitors: a review. Bioorg Med Chem, 17 (2): 444-59. [PMID:19095454]
3. Georgiadis D, Yiotakis A. (2008) Specific targeting of metzincin family members with small-molecule inhibitors: progress toward a multifarious challenge. Bioorg Med Chem, 16 (19): 8781-94. [PMID:18790648]
4. Grobe N, Di Fulvio M, Kashkari N, Chodavarapu H, Somineni HK, Singh R, Elased KM. (2015) Functional and molecular evidence for expression of the renin angiotensin system and ADAM17-mediated ACE2 shedding in COS7 cells. Am J Physiol, Cell Physiol, 308 (9): C767-77. [PMID:25740155]
5. Hoettecke N, Ludwig A, Foro S, Schmidt B. (2010) Improved synthesis of ADAM10 inhibitor GI254023X. Neurodegener Dis, 7 (4): 232-8. [PMID:20197648]
6. Levin JI, Chen JM, Laakso LM, Du M, Schmid J, Xu W, Cummons T, Xu J, Jin G, Barone D et al.. (2006) Acetylenic TACE inhibitors. Part 3: Thiomorpholine sulfonamide hydroxamates. Bioorg Med Chem Lett, 16 (6): 1605-9. [PMID:16426848]
7. Moss ML, Sklair-Tavron L, Nudelman R. (2008) Drug insight: tumor necrosis factor-converting enzyme as a pharmaceutical target for rheumatoid arthritis. Nat Clin Pract Rheumatol, 4 (6): 300-9. [PMID:18414459]
8. Niu X, Umland S, Ingram R, Beyer BM, Liu YH, Sun J, Lundell D, Orth P. (2006) IK682, a tight binding inhibitor of TACE. Arch Biochem Biophys, 451 (1): 43-50. [PMID:16762314]
9. Oarhe CI, Dang V, Dang M, Nguyen H, Gopallawa I, Gewolb IH, Uhal BD. (2015) Hyperoxia downregulates angiotensin-converting enzyme-2 in human fetal lung fibroblasts. Pediatr Res, 77 (5): 656-62. [PMID:25665060]
10. Oh M, Im I, Lee YJ, Kim YH, Yoon JH, Park HG, Higashiyama S, Kim YC, Park WJ. (2004) Structure-based virtual screening and biological evaluation of potent and selective ADAM12 inhibitors. Bioorg Med Chem Lett, 14 (24): 6071-4. [PMID:15546732]
11. Peng L, Cook K, Xu L, Cheng L, Damschroder M, Gao C, Wu H, Dall'Acqua WF. (2016) Molecular basis for the mechanism of action of an anti-TACE antibody. MAbs, 8 (8): 1598-1605. [PMID:27610476]
12. Rios-Doria J, Sabol D, Chesebrough J, Stewart D, Xu L, Tammali R, Cheng L, Du Q, Schifferli K, Rothstein R et al.. (2015) A Monoclonal Antibody to ADAM17 Inhibits Tumor Growth by Inhibiting EGFR and Non-EGFR-Mediated Pathways. Mol Cancer Ther, 14 (7): 1637-49. [PMID:25948294]
13. Walls AC, Park YJ, Tortorici MA, Wall A, McGuire AT, Veesler D. (2020) Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell, 181 (2): 281-292.e6. [PMID:32155444]
14. Xue CB, Voss ME, Nelson DJ, Duan JJ, Cherney RJ, Jacobson IC, He X, Roderick J, Chen L, Corbett RL et al.. (2001) Design, synthesis, and structure-activity relationships of macrocyclic hydroxamic acids that inhibit tumor necrosis factor alpha release in vitro and in vivo. J Med Chem, 44 (16): 2636-60. [PMID:11472217]
How to cite this page
M12: Astacin/Adamalysin: ADAM17. Last modified on 31/03/2020. Accessed on 02/08/2026. IUPHAR/BPS Guide to PHARMACOLOGY, https://www.guidetoimmunopharmacology.org/GRAC/ObjectDisplayForward?objectId=1662.




![]()
 This work is licensed under a Creative Commons Attribution-ShareAlike 4.0 International License
This work is licensed under a Creative Commons Attribution-ShareAlike 4.0 International License